Research
Current Projects
Communication between the microbiome and the immune system
By innovatively integrating microbiology, immunology, and carbohydrate and lipid chemistry to study the interactions between molecules and metabolites of the microbiota, our group has identified both microbial molecules and mechanisms responsible for immune system modulation and development. We used the intestinal mutualist Bacteroides fragilis to identify genetic loci for the synthesis of eight polysaccharides, designated PSA–PSH; we discovered that polysaccharide A (PSA) is sufficient to stimulate systemic/mucosal immune maturation. We overturned a classic paradigm of T cell activation by establishing PSA as a prototypical zwitterionic (equally positively and negatively charged) polysaccharide whose endolysosomal oxidative processing and presentation via the MIIC pathway (a route previously thought to be unique for proteins) activate CD4+ T cells. We demonstrated that PSA functions by stimulating both innate (TLR2 and Dectin-1) and adaptive immunity and that it is sufficient for restoring Th1/Th2 balance in normally Th2-skewed germ-free mice. We showed that orally administered PSA can protect mice from experimental inflammatory bowel disease. In addition, we showed that PSA presented by plasmacytoid dendritic cells treats and prevents experimental inflammatory bowel disease and experimental autoimmune encephalomyelitis, eliciting IL-10 production by regulatory T cells. We defined the microbiota’s role in early establishment of iNKT cell numbers in colon and lung. Our group identified another immunomodulatory microbial molecule, glycosphingolipid Bf717, which confers lifelong resistance to experimental ulcerative colitis through immunoregulation of proinflammatory iNKT cell homeostasis. To date, PSA and Bf717 are the only known immunomodulatory molecules from the microbiota. Our studies of bacterial–immune system interactions have remarkable physiologic and therapeutic implications.
We have done extensive immunoprofiling of 63 members of the gut microbiota and have conducted microarray analysis of the impact of monocolonization in order to understand how individual microbes modulate the immune system of the host. Also, using click chemistry, we have pioneered methods of labeling specific microbial molecules and visualizing them in the host’s immune system.
Mazmanian S, Liu C, Tzianabos AO, Kasper DL. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 2005; 122:107-118.
Mazmanian SK, Round JL, Kasper DL. A microbial symbiosis factor prevents intestinal inflammatory disease. Nature 2008; 453:620-625.
Chung H, Pamp SJ, Hill JA, Surana NK, Edelman SM, Troy EB, Reading NC, Villablanca EJ, Wang S, Mora JR, Umesaki Y, Relman DA, Kasper DL. Gut immune maturation depends upon colonization with a host-specific microbiota. Cell 2012; 1459:1578-1593.
Olszak T, An D, Zeissig S, Vera MP, Glickman JN, Baron RM, Kasper DL*, Blumberg RS* (*co-senior authors). Microbial exposure during early life has persistent effects on tissue-associated iNKT cells and their function. Science 2012; 336:489-493.
An D, Oh SF, Olszak T, Neves JF, Avci F, Erturk-Hasdemir D, Lu X, Zeissig S, Blumberg R, Kasper DL. Sphingolipids from a symbiotic microbe regulate homeostasis of host intestinal natural killer T cells. Cell 2014: 156:123-132.
Geva-Zatorsky N, Alvarez D, Hudak JE, Reading NC, Erturk-Hasdemir D, Dasgupta S, von Andrian UH, Kasper DL. In vivo imaging and tracking of host–microbiota interactions via metabolic labeling of gut anaerobic bacteria. Nature Medicine 2015; 21:1091-1100.
Surana N, Kasper D. Moving beyond microbiome-wide associations to causal microbe identification. Nature 2017; 552:244-247.
Geva-Zatorsky N, Sefik E, Kua L, Pasman L, Tan TG, Ortiz-Lopez A, Yanortsang TB, Yang L, Jupp R, Mathis D, Benoist C, Kasper DL. Mining the human gut microbiota for immunomodulatory organisms. Cell 2017; 168:928-943.
Hudak JE, Alvarez D, Skelly A, von Adrian UH, Kasper DL. Illuminating vital surface molecules of symbionts in health and disease. Nature Microbiol 2017; 2:17099.


Figure 1. Living Bacteroides in the intestine of a live mouse in real time. Multiple specific molecules are stained with different dyes (Hudak, et al., Nature Microbiology, 2017).
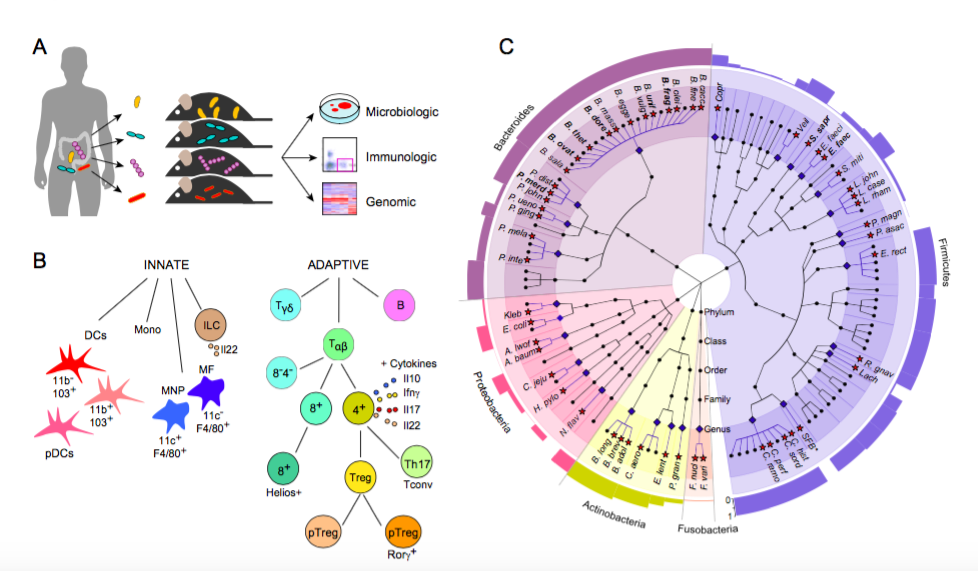
Figure 2. Immunoprofiling of host response to gut microbiota (Geva-Zatorsky, et al., Cell, 2017).
a) Experiment schematics: 4 week old germ free mice were mono-colonized with human gut microbes and analyzed after two weeks for microbial colonization, host immune response and host gene expression profile analyzed from colonic and small intestine whole tissue microarrays.
b) The immune cells analyzed in the study: Innate and adaptive immune responses were analyzed by flow cytometry of cells extracted from small intestines, Peyer’s Patches (PP), colons, mesenteric lymph nodes (MLN), and spleens of the mono-colonized mice. Innate cell types: Monocytes (Mono), Dendritic cells (DCs), Macrophages (MFs) and type 3 innate lymphoid cells (ILC3s). Adaptive cell types: B cells, gamma-delta T cells (Tγδ) and alpha-beta T cells (Tαβ), including cytokines (il10, il17, il22, IFNɣ).
c) Cladogram of the human gut microbiota generated by the GraPhlan software, [Huttenhower ref]. Microbes included in this cladogram are from the human microbiota project (HMP), 106 species detected by MetaPhlan, that were present in at least one subject at >0.5% abundance. Each phyla is represented by a different color, the genera that were included in this study are marked with blue diamonds and blue lines; the species covered in this study are marked with red stars (2-4 strains were analyzed for the species marked in bold, see table xx). The outer ring represents a bar graph of the prevalence of each genus, calculated by MetaPhlan1[ref, nat.meth.huttenhower], % genera with non-zero abundance. All species in this study were human associated microbes with the exclusion of Segmented filamentous bacteria (SFB, marked with a star), which is a mouse microbe.
Immunologic mechanisms of carbohydrate and glycoconjugate vaccine–induced immunity
Our research has established a new paradigm in the immunology of carbohydrates and polysaccharide–protein conjugates. Classically, carbohydrates were considered T cell–independent immunogens. We showed that one of the capsular polysaccharides from B. fragilis (PSA) is in fact able to induce an immune response (i.e., the induction of regulatory T cells) without a covalently attached protein. We demonstrated that the mechanism for this effect consists of processing (depolymerization by oxidative mechanisms) and presentation by antigen-presenting cells via the MHCII pathway. We showed that, in the endosome, the zwitterionic charge of PSA allows electrostatic binding interactions with MHCII and that this binding results in PSA presentation to the T cell receptor. We determined that plasmacytoid dendritic cells are critical for induction of IL-10 production by CD4+ T cells and that—in addition to presentation of the carbohydrate by MHCII—costimulation is required by CD86 and, ICOSL, which are in part stimulated by innate immunity through TLR2. We applied this concept to glycoconjugate vaccines. In the traditional model, a polysaccharide–protein conjugate was thought to be taken up by a polysaccharide-specific B cell, in which the carrier protein was degraded. A derivative peptide was bound to an MHCII molecule, which mediated its surface presentation to a T helper cell. Binding of the peptide–MHC complex to the T cell receptor elicited secretion of cytokines that stimulated the B cell to produce polysaccharide-specific antibodies. Through a series of studies, we showed that antigen-presenting cells internalize polysaccharide–protein conjugates and degrade the macromolecules to produce small peptide-glycan molecules, which bind to MHCII via the peptide moiety and are presented on the cell surface, where the glycan is recognized by carbohydrate-specific T cells. These observations radically changed our understanding of immune responses to carbohydrate antigens and yielded critical new insights invaluable in optimizing the design of glycoconjugate vaccines.
We have also worked for many years on creating glycoconjugate vaccines to prevent infection with Francisella tularensis. This organism presents interesting immunologic problems because immunity is directed against both an intra- and an extracellular phase of infection. We have created a number of F. tularensis mutants with altered lipopolysaccharide structures. This work has led us to understand that the lipopolysaccharide can be a critical mediator of immunity in both the intra- and extra-cellular phases of infection. We have now designed unique glycoconjugate vaccines that confer protection against challenge with this intracellular pathogen.
Cobb BA, Wang Q, Tzianabos AO, Kasper DL. Polysaccharide processing and presentation by the MHCII pathway. Cell 2004; 117:677-687.
Avci FY, Li X, Moriya T, Kasper DL. A mechanism for glycoconjugate vaccine activation of the adaptive immune system and its implications for vaccine design. Nature Medicine 2011; 17:1602-1609.
Sun X, Stefanetti G, Berti F, Kasper DL. Polysaccharide structure dictates mechanism of adaptive immune response to glycoconjugate vaccines. Proc Natl Acad Sci (USA) 2018; 116:193.
Stefanetti G, Okan N, Fink O, Gardner E, Kasper DL. A glycoconjugate vaccine using a genetically modified O antigen induces protective antibodies to Francisella tularensis. Proc Natl Acad Sci (USA) (in press, 2019).

Figure 1. Working model for PSA induced immunomodulation.

Figure 2. Mechanism of action for conjugate vaccines (Avci, et al., Nature Medicine, 2011).

Figure 3. Synthesis of a glycoconjugate vaccine.